In una remota regione dell'Australia settentrionale sono stati scoperte nuove specie di vertebrati con caratteristiche primitive. La scoperta si deve a ricercatori australiani dell'università "James Cook" in collaborazione con National Geographic Society. La spedizione, coordinata dal biologo Conrad Hoskin, lo scorso mese di Marzo si è recata in elicottero sulla catena montuosa di Cape Melville a Capo York, un complesso di roccia granitica all'estremo nord-est dell'Australia. Qui si trova una foresta pluviale incontaminata ed è proprio al suo interno che gli studiosi hanno scoperto 3 nuove specie di vertebrati sconosciute al mondo scientifico fino ad allora. Queste nuove specie sono state classificate dai tassonomisti; esse sono: un geco dotato di grandi capacità mimetiche (Saltuarius eximius) lungo circa una ventina di centimetri e con la particolarità di una coda che termina appiattita, come fosse una foglia. Inoltre questo geco è un animale prettamente notturno con occhi globosi molto ampi.
EPA/Conrad Hoskins / James Cook Univ /LANDOV
È stata trovata anche una rana maculata (Cophixalus petrolhilus) che vive a strettamente connessa nelle fessure del complesso granitico della catena montuosa di cape Melville; essa emerge solamente per nutrirsi e quando l'ambiente lo permette ovvero quando la stagione è più umida e piovosa.
EPA/Conrad Hoskins / James Cook Univ /LANDOV
Infine è stato individuato uno scincide, chiamatoSaprocincus saltus, un rettile dal color oro e con arti molto allungati; probabilmente adatti per vivere nelle insenature di rocce e terreno.
EPA/Conrad Hoskins / James Cook Univ /LANDOV
Questi animali possiedono tutti delle caratteristiche anatomiche e biologiche estremamente interessanti dovute all'isolamento territoriale che ha permesso loro di adattarsi all'ambiente circostante in maniera autonoma e senza ibridazioni con eventuali specie affini. Tra gli adattamenti riscontrati dall'equipe di ricercatori vi è quella della rana i cui girini si sviluppano all'interno delle uova in esemplari adulti in miniatura. Le uova vengono rilasciate, nella stagione secca, all'interno del granito in zone umide in modo che possano socchiudersi in totale sicurezza.
Questa è stata solo la prima di altre numerose spedizioni in questo "mondo perduto" dove si pensa di trovare ancora moltissime nuove specie tra cui molti invertebrati e si pensa possano anche esserci altri vertebrati... magari mammiferi.
In questi giorni d'autunno insolitamente caldi, sono state molte le segnalazioni di numerose coccinelle raggruppate su muri, tensostrutture ecc ecc. Quello che per molti si è rilevato un comportamento insolito di questi piccoli animaletti in realtà è un normale comportamento, tipico soprattutto della coccinella asiatica, Harmoniaaxyridis. Questa coccinella ,detta comunemente "coccinella cinese" oppure "coccinella Arlecchino"è un coleottero di origine asiatica temuto per la sua pericolosità in viticoltura e per il benessere umano.
Esso infatti nel periodo di vendemmia è solito penetrare all'interno dei grappoli per poi rilasciare la sua emolinfa. L'emolinfa di questa coccinella ha un odore sgradevole anche per l'uomo, dovuto a sostanze quali le Metossipirazine, che producono un odore di tipo "vegetale" simile a quello dei piselli e degli asparagi.
Le Harmoniaaxyridis furono introdotte in Europa e negli Stati Uniti per effettuare lotta biologica contro gli afidi che attaccano le piante (viti soprattutto) ma presto se ne perse il controllo sia negli U.S.A. (primo paese ad utilizzarle) ed in seguito in Ucraina e, da lì, nel resto dell'Europa. Ora sembra che in italia siano presenti in quasi tutto il nord fino alla Toscana.
Come dicevo all'inizio, non è insolito trovare aggregati di queste coccinelle poiché esse hanno l'abitudine di riunirsi in gruppi numerosi e compatti quando ci sono gli ultimi caldi estivi e, quindi, sono costrette a trovare luoghi dove "ibernarsi" per l'inverno. Solitamente la formazione di questo aggregato di coccinelle avviene attraverso l'arrivo di una piccola parte di coccinelle che potremmo definire "pioniere"; queste, trovato un posto adeguato, si stabiliscono nella zona e attraverso il rilascio di feromoni mandano un segnale di richiamo per le altre che giungono nel luogo da esse segnalato. E così, le coccinelle giunte sul luogo scelto, a loro volta emaneranno feromoni per richiamarne altre.
Solitamente prediligono luoghi d'aggregazione caldi e luminosi, per cui non è difficile trovarle attaccate a ombrelloni, gazebo e tensostrutture di colore chiaro; possono arrivare ad esserci anche 1 milione o più di Harmoniaaxyridis nel nascondiglio da loro scelto per l'ibernazione che spesso è un muro o un altro materiale inerte da costruzione.
Queste coccinelle sono un problema per l'uomo sia per le invasioni che possono produrre nelle abitazioni, sia perché posseggono degli allergeni che, in soggetti sensibili, possono indurre riniti allergiche, orticarie, asma e congiuntivite. Inoltre, durante lo svernamento, queste si trasferiscono dalle piante alle abitazioni (cercando il calore), incanalandosi nei traforati, nelle fessure ed in ogni insenatura possibile.
Qui possono rilasciare la loro emolinfa che, se è all'esterno, macchia visibilmente gli edifici; se invece esse si nascondono nei mobili, queste danneggiano l'arredo ed eventualmente anche vestiti e tendaggi.
Concludendo, non bisogna sottovalutare questo coleottero perché può risultare invasivo e creare anche problemi alla salute umana ma nemmeno allarmarsi troppo poichè nella maggior parte dei casi non si verificano invasioni massicce e pericolose.
In questi giorni si sono avute segnalazioni di aggregati di Harmoniaaxyridis in Lombardia, Emilia Romagna, Veneto e Toscana. Non abbiate paura di queste insetti ma tenete monitorata la situazione nell'eventualità che si avesse una massiccia invasione la prossima estate o le successive.
Chi ha visto queste coccinelle può stare tranquillo se non ha già avuto danni in casa o sulla propria pelle; anzi, la loro massiccia presenza è un segnale che questo caldo sta per finire e, come direbbe George R.R. Martin, "winter is coming" :-D
Harmoniaaxyridis
PER SAPERNE DI PIU': http://www.agrinotizie.com/articoli/news.php?id=1167#.Um6fvhAnOWM
Un particolarissimo e curioso squalo vive nelle profondità marine; è lo "squalo goblin" (o squalo folletto), conosciuto dai giapponesi fin dall'antichità come "Tengu-zame" e descritto per la prima volta dal mondo scientifico nel 1898 con il nome scientifico di: "Mitsukurina owstoni".
Il nome del genere (Mitsukurina) deriva da quello dello zoologo
giapponese Kakichi Mitsukuri, che diede il primo esemplare conosciuto a
D.Jordan per la descrizione e identificazione. Il nome
della specie (owstoni) invece viene da A.Owston, un collezionista
che acquistò il primo esemplare da un pescatore giapponese.
Questo squalo è l'unico membro vivente della famiglia dei Mitsukurinidae, ed è uno squalo abissale che nuota a profondità variabili tra i 300 ed i 1400 metri e le sue abitudini in alto mare sono ancora oggi poco conosciute per la difficoltà nel seguirlo e trovarlo nell'immensità degli oceani.
Lo squalo goblin può raggiungere i 3,3 metri di lunghezza e i 159 chili di peso ma spesso sono stati rinvenuti esemplari più piccoli sui 300-400cm.
La principale caratteristica che distingue questo animale è la stranissima forma della testa; lo squalo goblin possiede un lungo rostro simile ad un becco appiattito che è molto più lungo del muso delle altre specie di squalo.
Altra particolarità di questa specie è il colore del corpo, quasi completamente rosa-grigio, e le lunghe mascelle protrusibili grazie al fatto che la mascella non è fusa al cranio ma "in sospensione" attraverso legamenti e cartilagini. Quando la mascella viene estroflessa, in realtà i legamenti si rilassano, e si contraggono in seguito per far rientrare la mascella all'interno del cranio.
La rapida espansione della mascella provoca, inoltre, una sorta di vortice che aiuta lo squalo durante la predazione.
Secondo alcuni ricercatori, l'ampiezza d'apertura della mascella dello squalo goblin e la capacità così ampia di estrofletterla, è un adattamento ad un ambiente povero di scelte per quanto riguarda le possibili prede. Un adattamento di questo tipo permette a questo squalo di nutrirsi facilmente di prede delle dimensioni più diverse; dal piccolo crostaceo al grande cefalopode.
I suoi denti anteriori sono lunghi, lisci e sottili per cui sono abbastanza fragili soprattutto se paragonati a quelli posteriori che sono tozzi. La disposizione dentale fa pensare che le sue prede siano soprattutto piccoli pesci, cefalopodi e crostacei, prede catturate e trattenute dai denti anteriori e poi frantumate da quelli posteriori.
Quando le mascelle sono retratte all'interno della bocca, lo squalo goblin ricorda uno squalo toro (Carcharias taurus) madi colore rosa e con un naso insolitamente lungo.
La particolare colorazione rosa-grigio è dovuta non a pigmenti ma ad una pelle molto sottile ed insolita, se confrontata con gli altri squali, che lascia intravedere i capillari sotto cutanei. Questa pelle è molto sottile e molle e per questo molto delicata e facilmente soggetta a lacerazioni.
Le pinne hanno una
colorazione bluastra (dovuta alla grande capillarizzazione), le pettorali sono corte e larghe, le due dorsali
arrotondate e più piccole delle ventrali e della pinna anale, anche
questa arrotondata.
Estroflessione della mascella da parte di Squalo goblin
Lo squalo goblin deve cacciare in un ambiente dove l'oscurità la fa da padrone per cui non è sempre semplice individuare le poche prede che si trovano in profondità. Oltre ad utilizzare i suoi sensi si può notare che, se guardiamo lo squalo goblin dall'alto, si possono individuare gli occhi sulla cima del capo. Si pensa infatti che sia in grado di alzare gli occhi verso l'alto in modo da osservare le prede sopra di lui ed utilizzare così la poca luce che passa attraverso le profondità marine. L'occhio dello squalo goblin non possiede la membrana nittitante tipica di molti squali; gli squali che non la posseggono sono, infatti, solitamente capace di muover gli occhi all'indietro (come lo squalo bianco).
Altro adattamento che utilizza per individuare la preda sono delle strutture, chiamate ampolle di Lorenzini, presenti sul rostro. Queste strutture sono degli elettro recettori altamente sensibili che lo aiutano ad individuare il campo magnetico delle prede.
La conformazione corporea e gli adattamenti di questo squali fanno pensare che solitamente si muova in prossimità dei fondali marini, con spostamenti brevi e lenti.
La maggior parte degli esemplari di Mitsukurina owstoni catturati proviene dal Giappone prevalentemente in un'area compresa tra la Baia di Tosa e la Penisola di Boso. L'areale pacifico della specie è piuttosto esteso. Esemplari di M. owstoni sono stati trovati nelle acque al largo del Sudafrica e in vari siti sparsi un po' in tutto l'Oceano Pacifico occidentale. Altri squali goblin sono stati catturati al largo delle coste di Australia e Nuova Zelanda.
Nell'Oceano Atlantico la presenza della specie è stata confermata al largo della Guiana Francese a ovest e nel Golfo di Biscaglia e al largo di Madeira e del Portogallo a est. Dall'altro lato dell'Atlantico altri esemplari sono stati catturati anche nel Golfo del Messico.
Distribuzione geografica di Mitsukurina owstoni
Della sua riproduzione non si sa nulla ma si pensa siano ovovivipari come tutti i membri dell'ordine dei Lamniformi. Sebbene non sia mai stata catturata o avvistata una femmina gravida, si presume che le uova maturino e si schiudano all'interno del corpo della madre e i piccoli vengano partoriti vivi.
Recentemente ci si domanda come un animale così adattato alle profondità marine sia, in realtà, stato catturato molto spesso a profondità relativamente basse. Alcuni pensano sia uno squalo che vaga per le profondità alla ricerca di cibo e, quando il cibo scarseggia, si sposta verso la superficie rischiando in questo modo da essere catturato dai pescatori. Tuttavia non è considerato dalla IUCN (Unione Internazionale per la Conservazione della Natura), un animale a rischio estinzione poichè le sue normali abitudini abissali e le rare (e casuali) catture da parte dei pescatori, fanno pensare che non sia a rischio diretto. Infatti c'è sempre il rischio che, con il ridursi continuo e progressivo delle specie marine, lo squalo goblin possa rischiare la fame e, forse, questi primi avvistamenti in acque meno profonde possono essere un primo segnale di un declino trofico del sua areale.
Solo la ricerca scientifica ci potrà dare risposte a riguardo, nella speranza di non perdere, nel frattempo un organismo talmente affascinante e spettacolare impoverendo così i nostri mari di un altro "miracolo" della natura.
Per saperne di più:
- http://it.wikipedia.org/wiki/Mitsukurina_owstoni
- http://oubliettemagazine.com/2013/06/29/lincredibile-squalo-goblin-il-corpo-e-rosa-e-ricorda-una-sagoma-di-cartapesta/
- http://www.prionace.it/squalogoblin.htm#Nota%201
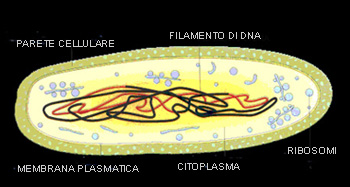
Negli anni settanta del XVII secolo, l’olandese Antony van Leeuwenhoek, mercante e fabbricante di lenti, osservò e descrisse per la prima volta gli organismi unicellulari simili ad animali che oggi vengono chiamati protozoi. Da allora, sono state descritte circa 35000 specie fossili e circa 30000 specie viventi. Ma cosa sono i protozoi? I protozoi sono organismi molto semplici sotto ogni punto di vista; la maggior parte di loro conduce vita libera e sono presenti in tutto il mondo ed in tutti i tipi di ambienti acquatici, compreso il terreno umido. Molte specie di protozoi sono parassite, e alcune delle malattie più dannose nel mondo sono provocate da protozoi. Come membri del regno protista (gruppo eterogeneo e polifiletico di organismi, che comprendono quegli eucarioti che non sono considerati né animali né piante o funghi), i protozoi differiscono dagli organismi pluricellulari in quanto tutti i loro processi vitali si svolgono all’interno di una singola cellula (organismi unicellulari). La diversità dei protozoi deriva principalmente dalle specializzazioni nelle membrane e negli organuli cellulari; organi caratteristici dei protozoi sono i vacuoli alimentari (funzione digestiva) ed i vacuoli pulsanti, con funzione di escrezione. Fig.1:Schema di un protozoo generico (Paramecium)
Alcuni protozoi sono coloniali, e formano così aggregati di cellule pressoché identiche tra loro, ma nessun protozoo possiede tessuti.
Inoltre, poiché i protozoi sono piccoli ed il loro rapporto superficie/volume è elevato, possono effettuare scambi gassosi e circolazione delle sostanze senza l’utilizzo di organuli specializzati.
La circolazione, solitamente avviene per diffusione intracellulare di sostanze disciolte ed i rifiuti azotati vengono eliminati sotto forma di ammoniaca che diffonde facilmente attraverso la membrana cellulare.
Frequente è l' incistamento, cioè l' eliminazione di buona parte dell'acqua citoplasmatica e il rivestimento di un involucro protettivo, denominato cisti, che resiste alla siccità e alle condizioni atmosferiche avverse. L'incistamento consente ai protozoi di diffondersi ampiamente, resistendo a situazione ambientali che per loro risulterebbero estreme.
Il fenomeno dell’incistamento è piuttosto raro nelle forme marine.
Generalmente i protozoi sono caratterizzati da forme ben definite (ad eccezione delle amebe e alcuni rizopodi), con diverse simmetrie (raggiata, sferica o bilaterale). Le loro dimensioni variano da circa 0.05mm a circa 5mm e soltanto dopo l’introduzione del microscopio elettronico è stato possibile osservare molta della diversità strutturale e della complessità dei protozoi.
MORFOLOGIA Membrana plasmatica: delimita il corpo cellulare dell’organismo. La struttura più diffusa è caratterizzata da un “mosaico fluido proteico” avvolto da un doppio strato lipidico. La membrana plasmatica ha la funzione principale di assumere sostanze nutritive, attraverso osmosi, fagocitosi, pinocitosi, citostomi … e di eliminare le scorie (escrezione) attraverso diffusione, esocitosi o attraverso il citopigio (apertura tipica di alcuni protozoi con funziona escretoria).
Glicocalice: È la parte più esterna della membrana plasmatica con funzioni di: - Barriera chimica e meccanica
- Interazione ospite-parassita
- Riconoscimento e ancoraggio al substrato ed a particolari strutture - Proprietà antigeniche Parete cistica: Si forma quando un protozoo passa dalla forma vegetativa a quella di cisti. La cisti è una forma di resistenza a fattori ambientali sfavorevoli. Questa parete protegge il protozoo dall’ambiente esterno e si origina dal rafforzamento esterno della membrana plasmatica grazie alla secrezione e al riassetto di particolari molecole (fosfoproteine, lipoproteine). Citoplasma: è la parte interna del protozoo che contiene numerose strutture e vari organuli Il citoplasma contiene: - Citoscheletro: è un complesso di elementi contrattili (filamenti di actina, miosina), organizzati in microtubuli, che sono responsabili del flusso citoplasmatico e, nelle amebe, del loro movimento “ameboide” - Mitocondri: sono presenti nei protozoi aerobi, sono simili a quelli degli organismi più evoluti (metazoi). La maggior parte dei protozoi possiede un unico lungo mitocondrio contenente un cinetoplasto, che è un organello all’esterno del nucleo a forma di bastoncello, ricco di dna, che governa la funzione respiratoria. - Idrogenosomi: presenti nei protozoi anaerobi; sono considerati analoghi ai mitocondri dal punto di vista funzionale ma lavorano in condizioni di anaerobiosi. - Reticolo endoplasmatico: ricco di ribosomi - Apparato di Golgi: non è presente in tutti i protozoi. - Lisosomi: contengono enzimi litici. Nucleo: contiene il DNA dell’organismo organizzato in cromosomi in numero e forma variabili. Sono presenti nucleoli contenenti RNA e non è raro trovare cellule polinucleate (con più nuclei). I protozoi ciliati possiedono un micronucleo (funzione riproduttiva) ed un macronucleo (funzione trofica).
Fig2: Altra immagine semplificata della struttura di un protozoo generico
MOVIMENTO
La locomozione può avvenire grazie alla presenza di strutture quali ciglia (generalmente molto numerose e corte) o flagelli (lunghi e presenti da uno a 8, a seconda dell’organismo). Particolare tipologia di locomozione è quella delle amebe, che sfruttano il movimento degli pseudopodi (=l'estroflessione mobile del citoplasma di un organismo unicellulare) per muoversi col tipico “movimento ameboide” che li caratterizza. Ciglia e flagelli possono anche avere funzione, nella maggior parte dei casi, sensoriale.
RIPRODUZIONE
I protozoi presentano svariate modalità di riproduzione. Tutti sono in grado di riprodursi asessualmente per scissione binaria, per scissione multipla o per gemmazione; le cellule figlie rigenerano eventualmente gli organuli non ripartiti equamente; Alcuni protozoi si riproducono esclusivamente per via asessuale. La riproduzione sessuale è anch’essa molto comune e prevede sia la formazione di isogameti sia di anisogameti, questi ultimi anche ben differenziati come spermi e uova; nei Ciliati la riproduzione sessuale prende la forma di coniugazione, che non comporta la differenziazione di gameti; più rara è l'autogamia, ovvero L'unione di gameti, maschile e femminile, provenienti dallo stesso individuo (organismo ermafrodito). Forme cistiche si possono formare anche intorno agli zigoti o intorno a individui prima che avvengano i vari processi riproduttivi. La formazione delle cisti ha un ruolo importante, ma non esclusivo, anche nella dispersione dei protozoi nell’ambiente esterno poiché ne conferiscono resistenza all’ambiente favorendone così il trasporto anche lontano. Classificazione La classificazione dei Protozoi non è semplice ed è continuamente soggetta a modificazioni da parte degli specialisti, poiché le nuove tecnologie e l’utilizzo della biologia molecolare permettono di classificare in maniera differente e più precisa gli organismi viventi. Per cui utilizzerò una delle tante classificazioni possibili, quella che considero più semplice e sicuramente utile anche a chi non è strettamente connesso col mondo scientifico e della sistematica. Ci sarebbero 6 phyla di protozoi di cui 4 sono quelli più numerosi e di interesse per l’uomo. PHYLUM SARCOMASTOGOPHORA (Sarcomastigofori) Questo phylum comprende tre tipi di protozoi. Quelli che si muovono per mezzo di prolungamenti citoplasmatici detti pseudopodi sono detti amebe; quelli che hanno uno o più flagelli sono detti flagellati; mentre gli ameboflagellati hanno sia pseudopodi che flagelli durante differenti stadi del ciclo biologico. L’esistenza degli ameboflagellati indica che le amebe ed i flagellati sono strettamente affini ed è questa la ragione per cui questi organismi vengono inseriti nello stesso phylum. Amebe: Le specie del genere Amoeba sono molto comuni sul fondo degli stagni e dei laghi d’acqua dolce dove si muovono e si alimentano mediante l’azione degli pseudopodi. Il loro ciclo biologico comprende uno stadio di cisti dormiente che permette loro di sopravvivere ai periodi di siccità mentre durante lo stadio di ameba “attiva” detto trofozoite, si riproducono per via asessuale (scissione binaria). Molte amebe sono rivestite da una singola membrana cellulare detta plasma lemma ma alcune amebe possiedono anche un involucro esterno detto guscio o teca che circonda e protegge il plasma lemma (Es. Arcella e Chlamydophrys). Il guscio è formato da secreti proteici induriti e talvolta rivestiti da granuli di sabbia o scaglie silicee. Gli pseudopodi vendono chiamati lobopodi se sono larghi, a forma di vanga o tubolari (Es. Arcella) mentre sono detti filopodi quando sono filiformi e sottili verso le estremità (Es. Chlamydophrys). Fig3:Esempio di ameba (Amoeba proteus)
I foraminiferi, un grande gruppo di amebe marine, sono molto importanti ecologicamente. La maggior parte delle specie secerne un guscio (nicchio) di carbonato di calcio nel quale l’ameba vive.
Per muoversi utilizzano dei prolungamenti cellulari molto ramificati detti reticulopodi (o rizopodi o anche mixopodi). Questo reticolo permette ai foraminiferi di strisciare lentamente sul fondo del mare; inoltre questo reticolo è rivestito da un muco appiccicoso che viene utilizzato per catturate microrganismi usati come alimento. Si stima che 1/3 del fondo degli oceani e dei mari del mondo sia ricoperto di spessi depositi di gusci di carbonato di calcio provenienti da foraminiferi fossili. Le bianche scogliere di Dover sono costituite da foraminiferi fossili. Fig.4:Foraminiferi
Un altro gruppo di amebe sono gli attinopodi, detti anche radiolari o eliozoi. Sono protozoi sia d‘acqua dolce che marini i quali secernono scheletri complessi. Gli attinopodi hanno pseudopodi simili ad aculei, molto sottili alle estremità, detti assopodi. Questi s’irraggiano dal corpo cellulare centrale sferico. La forma sferica degli attinopodi è sostenuta da uno scheletro interno. Gli attinopodi si muovono rotolando lentamente qua e là come plancton oppure sul fondo di laghi, oceani e mari retraendo e allungando gli assopodi.
Fig.5:Radiolari (attinopodi)
Molte amebe sono parassite e si trasferiscono da un ospite all’altro come cisti dormienti. Entamoeba histolytica comprende uno stadio ameboide attivo che vive nell’intestino umano ed una cisti sferica che viene espulsa attraverso le feci. La malattia è più frequente nelle zone tropicali e, in generale, dove le condizioni sanitarie sono mediocri. I soggetti s’infettano ingerendo la cisti.
Flagellati: Sono rivestiti da una pellicola, un involucro composto da una o più membrane cellulari e frequentemente rinforzato da microtubuli sottostanti. Questo rivestimento rende i flagellati meno flessibili delle amebe e, mentre gli pseudopodi posso sporgere da qualsiasi regione di un ameba, i flagellati conservano tipicamente una forma particolare. Euglena gracilis, comune negli stagni d’acqua dolce, ha tipicamente una forma allungata con una estremità anteriore arrotondata ed estremità posteriore assottigliata. Molti flagellati sono autotrofi simili a piante e contengono granuli verdi, giallastri o rossi, detti plastidi. Fig.6:Euglena gracilis Altri invece sono eterotrofi, incolori e più simili ad animali. Alcuni però sono autotrofi, ma sono anche capaci di nutrizione eterotrofica. Un esempio di quest’ultima particolarità possiamo a verla coltivando in laboratorio colture di E. gracilis; si è notato che, se le si coltivano nell’oscurità, perdono la propria capacità fotosintetica (perdono i plastidi) e appaiono di una colorazione molto neutra. Se a questi nuovi individui, privi di attività fotosintetica, vengono fornite sostanze nutritive organiche disciolte, gli individui possono sopravvivere e riprodursi per anni come eterotrofi. Inoltre, se tali individui vengono esposti alla luce, compaiono cloroplasti attivi e gli organismi possono ridiventare autotrofi. Molti flagellati sono parassiti. Le specie di Trypanosoma, per esempio, infettano l’apparato circolatorio di molti vertebrati. La tripanosomiasi africana (o malattia del sonno dell’Uganda) e la malattia di Chagas (tripanosomiasi americana o brasiliana o sudamericana) sono causate da tripanosomi che infettano l’uomo attraverso organismi vettore o per contatto diretto in ambienti acquatici ricchi di questi flagellati. Anche l’economia in alcune regioni africana è stata compromessa da alcune tripanosomiasi; in particolare da una tripanosomiasi detta “Nagana” la quale uccide bovini, equini ed altri animali d’allevamento. La tripanosomiasi africana e il Nagana sono trasmessi da mosche tse-tse, insetti ematofagi che possono trasmettere gli stati infettivi dei parassiti. La malattia di Chagas viene trasmessa all’uomo da insetti ematofagi della famiglia dei “reduvidi” (reduviidae).
Fig.7:Esempio di Trypanosoma Euglena e Trypanosoma hanno un solo flagello mentre molti flagellati hanno 2 o più flagelli; le specie di Trychonympha, che vivono nell’intestino delle termiti e delle blatte xilofaghe e digeriscono la cellulosa contenuta nel legno ingerito dagli ospiti, hanno molti flagelli.
Fig.8:esempio diTrychonympha Molti flagellati sono aggregati coloniali costituiti da un piccolo numero a molte migliaia di cellule. Le colonie si sviluppano nel momento in cui le cellule si dividono, rimanendo però attaccate fra loro. Le specie di Volvox sono autotrofi coloniali, costituiti da colonie di colore giallo o verde, talmente grandi da essere visibili ad occhi nudo. Le colonie di Volvox sono comuni negli habitat d’acqua dolce poco profonda, si muovano ruotando lentamente grazie alla forza combinata di migliaia di flagelli vibranti (due su ciascuna delle cellule della colonia).
Un altro gruppo di flagellati interessante sono i coanoflagellati. Possono essere sia cellule solitarie che colonie e le loro cellule sono tipicamente peduncolate ed hanno un collare trasparente che circonda un singolo flagello. Il flagello, con la sua azione, convoglia verso il collare le particelle in sospensione nell’acqua. Il collare può agire come un setaccio, catturando piccole particelle che vengono ingerite dal copro cellulare.
Per struttura e modo di alimentarsi i coanoflagellati somigliano strettamente ai coanociti (cellule con collare) dei poriferi (spugne) e molti zoologi ritengono che i poriferi si siano originati da protozoi simili ai coanoflagellati. Fig.10: Coanoflagellati (singola cellula e colonia)
Ameboflagellati: Tra questi organismi, amebe dotate di flagelli, i più comuni sono le specie del genere Naegleria, che abitano il terreno umido e gli ambienti d’acqua dolce ricchi di sostanza organica. Talvolta possono trovarsi sul fondo di piscine non adeguatamente igienizzate. Una specie, Naegleria fowleri, può causare una malattia umana letale, la meningoencefalite amebica se riesce a raggiungere l’encefalo attraverso le narici. Le specie di Naegleria sono tipicamente ameboidi in condizioni favorevoli, ma si possono sviluppare stadi flagellati quando scarseggiano le sostanze nutritive. I ricercatori ipotizzano che i flagelli permettano a Naegleria di allontanarsi nuotando dagli habitat sfavorevoli e di dirigersi verso quelli favorevoli. PHYLUM APICOMPLEXA (Apicomplessi) Questo phylum deve il suo nome ad un complesso di organuli situati all’estremità assottigliata, o apice, della cellula. Tutti gli Apicomplexa sono parassiti, e gli organuli apicali permettono loro, probabilmente, di penetrare attraverso le membrane cellulari o gli strati di tessuti de loro ospiti. Fig.11: Apicomplexa generico
Il ciclo biologico della maggior parte degli Apicomplexa comprende la moltiplicazione asessuale per scissione multipla (schizogonia), una fase sessuale detta gametogonia, ed una fase asessuale detta sporogonia. La maggior parte degli Apicomplexa appartiene alla classe Sporozoea (sporozoi), che comprende due sottoclassi Gregarine (parassiti extracellulari che abitano il cavo orale ed il canale alimentare di molti invertebrati), e Coccidia (coccidi) che sono spesso parassiti intracellulari di molti animali domestici che causano importanti malattie (Toxoplasma gondii, generalmente innocuo per un umano adulto ma molto dannoso per un feto).
I coccidi più importanti dal punto di vista delle patologia umane sono sicuramente le specie appartenenti al genere Plasmodium, tra cui troviamo gli organismi che causano la malaria. Il ciclo biologico dei Plasmodium è molto complesso ed implica riproduzione asessuale, che avviene con modalità differente negli ospiti di questo protozoo) ed una fase sessuale con stadi in entrambi gli ospiti.
PHYLUM MICROSPORA (Microsporei) I Microsporei sono parassiti intracellulari che formano spore particolari formate da un corpo centrale di sporoplasma ed un filamento cavo ( tubo) convoluto il quale si estroflette quando la spora viene assimilata dall’ospite e lo sporoplasma viene iniettato nell’ospite stesso attraverso il tubo che nel frattempo si estroflette. Come gruppo, infettano un’ampia varietà di animali nella maggior parte dei phyla, e molte specie sono “ospite-specifiche” ovvero infettano un’unica specie ospite. Lo stadio infettivo dei Microsporei è dato dallo stadio di spora, le quali sono minuscole (5µm). Particolarità di queste spore è che non possiedono mitocondri, per cui ricavano l’energia necessaria al loro accrescimento e sviluppo dall’ospite parassitato. Dal punto di vista economico-commerciale sono importanti le specie di Nosema che causano malattie nelle api domestiche e nei bachi da seta.
PHYLUM CILIOPHORA (ciliofori) I ciliati (ciliofori) rappresentano il phylum più numeroso di protozoi e sono caratterizzati dalle ciglia e da un processo riproduttivo sessuale detto coniugazione. Possiedono, inoltre, una pellicola pluristratificata sostenuta da fibrille sottostanti, e da due differenti tipi di nuclei; un macronucleo ed uno o più micronuclei. Il macronucleo dei ciliati e tipicamente poliploide (cioè ha multipli del numero diploide di cromosomi: 2, 4, 6….2*n) ed ha il compito di regolare l’alimentazione, gli scambi gassosi, l’osmoregolazione e altre funzioni non-riproduttive. I micronuclei invece sono diploidi e subiscono la meiosi prima della coniugazione. Durante la coniugazione, vengono scambiati micronuclei aploidi tra gli individui (ciò fornisce variabilità genetica al novo individuo in formazione). I ciliati sono presenti frequentemente in gran numero in pozze stagnanti d’acqua dolce o salata, nonché nel terreno umido. Parecchie specie sono mantenute in coltura e sono diventate organismi importanti nella ricerca come, ad esempio, alcune specie di Paramecium di cui si sono interessati biologi cellulari, genetisti ed ecologi scrivendo anche molti libri a riguardo.
Le cellule di Paramecium hanno una tipica forma “a ciabatta” e sono rivestite da ciglia e presentano anche una regione orale distinta (citostoma). Un altro ciliato, Didinium nasutum, è un predatore vorace di Paramecium che paralizza la sua preda prima di nutrirsene. Molto usato nella ricerca, in particolare in genetica ed in tossicologia, è il piccolo ciliato d’acqua dolce Tetrahymena pyriformis; questo ciliato è grande un quinto rispetto ad un Paramecium ed è a forma di lacrima. Come Paramecium si alimenta soprattutto di batteri e può essere mantenuto in coltura per ricerche scientifiche. Degno di nota, infine, èStentor coeruleus, considerato dagli addetti ai lavori molto bello per la sua colorazione blu. Solitamente è attaccato al substrato ma talvolta può essere trovato natante in ambiente acquatico. Habitat e ambiente dei protozoi I protozoi sono capaci di accrescersi e riprodursi solamente in habitat umidi poiché la loro membrana superficiale deve rimanere sempre umida per facilitare gli scambi gassosi e le altre funzioni vitali e si trovano praticamente ovunque ci sia acqua. Popolazioni dense compaiono spesso in stagni che si prosciugano stagionalmente ed in pozze che durano pochi giorni ed anche nelle schiume superficiali di acque stagnanti. Alcune specie si nutrono di plancton (in sospensione nell’acqua), mentre molte volte vivono attaccate a piante, animali e superfici non appartenenti ad organismi viventi. Nel terreno umido, i protozoi vivono in piccole goccioline e pellicole d’acqua e attorno a particelle di terreno, microhabitat troppo piccoli per la maggior parte degli animali. Le amebe, grazie al loro copro flessibile, manovrabile ed elastico, sono frequentemente abbondanti nei terreni umidi, insieme a molti piccoli ciliati e flagellati. La maggior parte dei protozoi terricoli hanno uno stadio di cisti dormiente in cui sopravvivono a periodi di gelo e siccità; quando poi il clima è più favorevole i protozoi emergono dalla loro forma di ciste per nutrirsi e riprodursi. La forma di ciste è molto utile anche perché permette la dispersione dei protozoi nell’ambiente attraverso il vento, le correnti d’acqua ed il contatto con animali. Ruolo nelle catene alimentariLe popolazioni di protozoi sono fondamentali nelle catene alimentari acquatiche. Alla base di una catena alimentare, la fotosintesi per opera dei flagellati verdi fornisce frequentemente molta energia organica disponibile negli ambienti d’acqua dolce e marini. Molte amebe, flagellati eterotrofi e ciliati sono predatori che si alimentano di batteri, diatomee, lieviti alghe ed altri protozoi. Altri protozoi sono definiti “scavenger” (spazzini), poiché si alimentano di sostanza organica morta. Come i batteri, gli spazzini aiutano a decomporre le piante e gli animali morti e a riciclare le sostanze nutritive riconvogliandole nelle catene alimentari. I batteri sono l’alimento più comune per i protozoi. Le specie che si alimentano di batteri, e a loro volta sono consumate da piccoli animali o da altri protozoi, formano un’importante connessione nutritiva fra i batteri decompositori e gli altri organismi delle catene alimentari. Le popolazioni di protozoi che si alimentano di batteri raggiungono molto spesso un ammontare enorme; lo stesso vale per quelle specie che sono capaci di svolgere la fotosintesi. Infatti, le “fioriture”, che sono grandi popolazioni monospecifiche (= formate da una sola specie) di certi flagellati autotrofi, si formano in laghi, oceani e mari quando le condizioni ottimali promuovono la riproduzione rapida. Le fioriture si possono presentare come vaste aree rossastre o verdastre sull’acqua. Le famose acque rosse (red tide), comuni lungo le coste orientali e occidentali degli Stati uniti, sono causate da fioriture di una quantità enorme di flagellati, tra cui Gymnodinium brevis. Le tossine prodotte da questi flagellati causano a volte la moria di pesci nelle zone di maggior concentrazione della tossina, e soggetti umani sono stati colpiti da paralisi o sono morti per aver ingerito prodotti pescati contaminati da queste tossine. I protozoi sono importanti anche dal punto di vista ecologico poiché presenti ovunque e perché parassitano organismi, nutrendosi di loro, a volte in maniera asintomatica altre in modo più invasivo. Si vengono così a creare degli ospiti che in realtà fungono da “nicchia ecologica” per questi piccoli organismi, antichi e temibili abitanti del nostro affascinante pianeta. Fig.14: Fenomeno del “Red tide”, dovuto alla fioritura di protozoi (in questo caso Lingulodinium polyedrum) Bibliografia e link utili: - Lawrence G. Michell et al. Zoologia, Zanichelli. 1999 - http://www.luciopesce.net/zoologia/protoz.html - http://it.wikipedia.org/wiki/Protozoa - http://www.amicidelmicroscopio.it/protozoi.php Tag di Technorati: scienza,protozoi,ciliati,flagellati,amebe,zoologia,paramecio
Immagine da:http://www.naturamediterraneo.com/forum/topic.asp?TOPIC_ID=8462
Nella mia città (Modena) ci sono piante curiose che sicuramente avete avuto modo di individuare durante le vostre passeggiate. Una di queste è sicuramente la Maclura pomifera, una pianta legnosa dal portamento arboreo della famiglia delle Moraceae (come il gelso per intenderci), conosciuta nella cultura popolare anche come “melo da siepe”o “Moro degli Osagi”.
È un albero dioico, ovvero esistono piante “maschili” e “femminili”, di altezza variabile tra i 6 e 20 metri. La corteccia di questa pianta è color bruno-arancio e contiene tannini, è rugosa ed è caratterizzata dalla presenza di molte fessure nelle piante più anziane. Vi sono inoltre spine lunghe e robuste sulle ascelle fogliari ed il legno è molto duro, compatto, resistente e flessibile. La caratteristica di questo legno ha fatto sì che questa pianta venisse usata, presso la tribù degli indiani d’America degli Osagi, come legno per la produzione di archi. Dalle radici di questa pianta si estrae un pigmento chiamato morina, estraibile anche da “parenti” di questo albero come il gelso e l’albero del mirto. La morina è un colorante giallo molto intenso, utilizzato per la colorazione di lana e cotone e veniva anche utilizzato mescolato ad altre sostanze (come l’indaco) per produrre altri coloranti. Le foglie sono molto simili a quelle dell’albero dell’arancio e sono caratterizzate dal possedere una parte superiore glabra ed una pagina inferiore pelosa. In passato queste foglie sono state utilizzate per nutrire i bachi da seta (Bombyx mori) ma l’introduzione di queste foglie non ebbe molto successo perché meno nutrienti rispetto alle foglie del gelso, normalmente utilizzate per l’alimentazione dei bachi. Questa infruttescenza è un ammasso sferico di colore variabile dal giallo al verde, di consistenza legnosa/coriacea e con la superficie profondamente corrugata. Il frutto aperto rivela una polpa biancastra da cui cola un liquido lattiginoso. Il frutto non è commestibile e se ingerito può provocare all’essere umano nausea e vomito (emetico) mentre è un frutto commestibile e ben tollerato dagli scoiattoli. Come precedentemente detto gli Osagi utilizzavano il legno di questa pianta per produrre archi molto resistenti ma anche come rimedio contro congiuntiviti ed infiammazioni oculari. Oggi questa pianta ha funzione ornamentale e per la realizzazione di siepi dall’aspetto impenetrabile, dovuto anche alle spine presenti, e decorative per merito delle sfumature del legno che vanno da un marrone scuro al color ocra. Alcune curiosità: Questo stesso composto viene utilizzata anche contro la Candida, micete parassita dell'uomo
Originariamente questa pianta era tipica del nord America, più precisamente in un area degli Stati Uniti centrali (Texas ed Arkansas) abitata dalla tribù, precedentemente citata, degli Osagi. Nel nord America questa pianta viene chiamata anche arancio degli Osagi (Osage orange) , proprio dal nome della tribù indiana che risiedeva nella zona di crescita di questo albero.
La specie è dioica, cioè ha fiori maschili e femminili su piante differenti ed i fiori sono riuniti in infiorescenze. La caratteristica più curiosa della pianta, e che sicuramente fa notare ai passanti la presenza di questo albero nei nostri parchi, è il frutto che è più precisamente una infruttescenza (sorosio) formata da un insieme di acheni ognuno derivante da un diverso ovario.
- Nell'ultima guerra mondiale la corteccia della Maclura pomifera è stata utilizzata per produrre il colore “Kaki” delle divise americane
- Il pigmento “morina” che si ricava da questa pianta, veniva utilizzato dagli Osagi per tingersi il volto.
- In un articolo pubblicato nel 2003 sulla rivista della Botanical Society of America, si discute della tossicità delle foglie nei confronti degli erbivori
- Alcuni studi affermano che la linfa di questa pianta possa provocare dermatiti nell’essere umano.
- In M. pomifera è stato individuato il Resveratrolo (3,5,4'-triidrossi-trans-stilbene), un alcaloide fungicida che spiegherebbe l’eccezionale resistenza del legno della Maclura alla degradazione fungina.
- In Kansas la pianta di Maclura viene detta anche “siepe”
- Ebbe grande diffusione negli Stati Uniti (prima in Illinois) perché veniva utilizzato per recintare i terreni agricoli in modo efficiente e poco costoso.
- Il legno della Maclura è fra i più pesanti d’America: un metro cubo allo stato naturale pesa più della metà di un metro cubo di calcare, ed è quasi altrettanto duro perché smussa rapidamente le punte da tornio e le lame da sega; inoltre, pur essendo prodigiosamente flessibile, è due volte e mezzo più resistente del legno di quercia: un arco di arancio Osage, fatto con una pianticella ben stagionata e flesso con un tendine di bufalo, può scagliare una freccia di corniolo con tanta forza da farla penetrare in un bisonte fino alle penne, e tutt’oggi alcuni arcieri considerano il suo legno superiore al celeberrimo tasso usato per gli archi inglesi. Nel 1811 John Bradbury, incoraggiato da Jefferson, fece un viaggio di esplorazione lungo il Missouri River e disse che il prezzo di un arco di Maclura era elevatissimo poiché ammontava ad un arco e ad una coperta.
- Il legno di Maclura veniva utilizzato dai coloni per produrre gli assali dei carri, i mozzi delle ruote, le pulegge, i manici degli attrezzi, i pali del telegrafo, gli isolatori, i manganelli e le traversine ferroviarie. Per saperne di più: http://www.racehorseherbal.com/Wild_Herbs/Osage_Orange/osage_orange.html http://www.actaplantarum.org/floraitaliae/viewtopic.php?t=11542 http://luirig.altervista.org/flora/taxa/index1.php?scientific-name=maclura+pomifera http://giardinaggioirregolare.com/2010/04/20/nel-reticolo-maclura-pomifera/ http://plants.usda.gov/core/profile?symbol=MAPO http://it.wikipedia.org/wiki/Maclura_pomifera
È un fenomeno che consiste nell’immissione luminosa da parte di organismi viventi attraverso particolari reazioni chimiche che convertono l’energia chimica in energia luminosa. La bioluminescenza è stata trovata in organismi quali molluschi, anellidi ctenofori, pesci, dinoflagellati, insetti e funghi.
Gli organismi marini, hanno organi appositi per la produzione di luminescenza che prendono il nome di fotofori. Questi si trovano nei pesci, nei cefalopodi e negli cnidari.
Gli scopi della bioluminescenza possono essere molteplici: A scopo difensivo, difesa, comunicazione, attacco, illuminazione…
La bioluminescenza può avvenire in 2 modi: 1) Bioluminescenza propria dell’organismo; dovuta a processi biochimici che avvengono nel organismo.(Es: lucciole, 2) Simbiosi con batteri bioluminescenti che utilizzano delle reazioni redox per produrre luminescenza.
Meccanismo della bioluminescenza
La bioluminescenza si basa su un meccanismo per cui alcune molecole, prodotte in uno stato elettronico eccitato, emettono parte di energia sotto forma di radiazione luminosa tornando allo stato fondamentale di quiete.
Diversi studi biochimici e biomolecolari hanno dimostrato che i meccanismi di emissione di luce da parte di organismi viventi sono molto vari e quindi, con buona probabilità, tale proprietà si è sviluppata in maniera indipendente nei vari gruppi biologici.
È stato inoltre accertato che in molti organismi viventi, tra i quali la lucciola, la bioluminescenza implica l'azione di almeno due composti chimici: un substrato organico che emette la luce, chiamato "luciferina", e un enzima catalizzatore chiamato "luciferasi". Nella maggior parte dei casi il fenomeno è appunto dovuto alla luciferina, che in presenza di ATP (adenosintrifosfato), magnesio e dell'enzima luciferasi, cede elettroni, i quali, passando ad un livello minore di energia, liberano energia sotto forma di luce.
Nei batteri (Es: Photobacterium) la bioluminescenza è associata al consumo di ossigeno (O2) e quindi avviene solo in condizioni di aerobiosi e di alte concentrazioni di questi organismi nell’ambiente.
In alcuni organismi la bioluminescenza è fatta da batteri in simbiosi mutualistica con l’organismo ospite a punto che i processi biochimici della luminescenza sono strettamente collegati ed inseparabili dall’organismo ospite. Ad esempio batteri luminescenti vivono in simbiosi con un pesce chiamato Photoblepharon palpebratus(Fig.1) della barriera corallina australiana e si sviluppano all’interno di una piega (palpebra) sotto agli occhi. La simbiosi fra questi organismi fa sì che il pesce fornisca un substrato protettivo e molto nutriente ai batteri ed i microrganismi offrano la luce necessaria per le diverse comunicazioni dell’animale atte a difendere il proprio territorio e a cacciare.
Un altro esempio di simbiosi fra batteri luminescenti e animali è fornita dall’interazione fra Euprymna scolopes (Fig,2), un cefalopode che di giorno vive sotto la sabbia e di notte vaga in cerca di cibo, e Vibrio fischeri. Euprymna riesce a mimetizzarsi nell’ambiente notturno grazie alla presenza di un organo luminescente (fotoforo) che emette una luce “lunare” che ne fa scomparire l’ombra. La luce è emessa da popolazioni di Vibrio fischeri, batteri bioluminescenti che vivono all’interno del fotoforo del cefalopode dove trovano un ambiente ricco di nutrimento.
La possibilità di emettere radiazioni luminose si trova in un ristretto numero di specie che hanno principalmente abitudini notturne notturne o che vivono in luoghi oscuri come, ad esempio, caverne o l’interno di alberi. La luminescenza può originarsi: - Perché l'insetto possiede speciali organi fotogeni - Come effetto collaterale del suo metabolismo - Perché causata da batteri patogeni e quindi essere sintomo di malattia (es. larve di Mamestra attaccate dal Bacterium haemosphoreum) - Perché ospita batteri luminescenti simbiontici.
Le specie luminose si riscontrano prevalentemente tra i coleotteri (elateridi, drilidi, lampiridi, fengoidi), ma anche fra i collemboli, i rincoti omotteri cicadoidei, e le larve dei ditteri. Nelle larve di alcuni ditteri fungivoridi la luce proviene dal corpo adiposo o dall'estremità ingrossata dei tubi malpighiani (strutture paragonabile ai reni dei vertebrati), nei coleotteri fengoidi da agglomerati di enociti (ghiandole ). Nei coleotteri, che hanno differenziato organi luminosi, può essere emessa luce in tutti gli stadi di sviluppo.
La funzione della bioluminescenza, nelle forme più primitive, sembra non avere alcun significato biologico, essendo un effetto collaterale del metabolismo; nelle larve luminose di alcuni ditteri fungivoridi essa probabilmente serve da attrattivo per le prede e nei coleotteri lampiridi, fengoidi ed elateridi è un evidente richiamo sessuale.



.jpg/600px-Asian_lady_beetle-(Harmonia-axyridis).jpg)






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}